



Quá trình apoptosis, phản ứng oxy hóa và cấu trúc phiên mã của tế bào niêm mạc dạ dày, qua đó nghiên cứu ảnh hưởng của Sulforaphane đến những quá trình này.
Thông tin nghiên cứu
Từ khóa:
- Quá trình apoptosis
- Isothiocyanate
- Chất chống oxy hóa
- Cytoprotective
- Dinh dưỡng
- Giải trình tự
Tổng quan
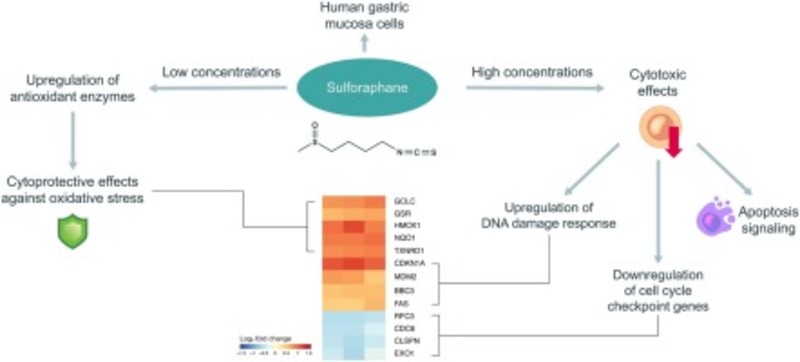
Stress oxy hóa là một yếu tố quan trọng trong cơ chế sinh bệnh của một số bệnh đường tiêu hóa. Sulforaphane (SFN), một hợp chất với hoạt tính sinh học mạnh được tìm thấy trong các loại rau họ cải được cho là có khả năng kích hoạt yếu tố 2 liên quan đến hồng cầu hạt nhân nhạy cảm với oxy hóa khử 2 (NRF2). Ngoài vai trò bảo vệ, SFN còn có tác dụng gây độc tế bào đối với tế bào ung thư. Tuy nhiên, thông tin liên quan đến độc tính của SFN trong tế bào bình thường vẫn còn thiếu khá nhiều. Nghiên cứu này sẽ đánh giá tác động của SFN đối với khả năng tồn tại của tế bào, khả năng chống oxy hóa và biểu hiện gen trong tế bào niêm mạc dạ dày của con người (MNP01). Quan sát cho thấy, SFN làm giảm sự hình thành ROS và bảo vệ tế bào chống lại stress oxy hóa gây ra. Tuy nhiên, nồng độ SFN cao sẽ làm tăng quá trình apoptosis. Nồng độ SFN trung gian (8 μM) được chọn cho các nghiên cứu giải trình tự RNA. Chúng tôi đã quan sát sự điều chỉnh các gen của con đường NRF2 (chất chống oxy hóa), phản ứng tổn thương DNA, và tín hiệu apoptosis; trong khi SFN điều hòa chu kỳ tế bào và gen sửa chữa DNA. Từ đó, nghiên cứu cho ra kết quả rằng SFN có thể bảo vệ tế bào ở nồng độ thấp và gây độc tế bào ở nồng độ cao.

1. Giới thiệu
Chế độ ăn “kiểu phương Tây”, đặc trưng bởi lượng đường tinh luyện và chất béo bão hòa cao, có liên quan đến béo phì và các bệnh khác, bao gồm cả ung thư đường tiêu hóa (GI). Ngoài ra, chế độ ăn giàu chất béo và ít chất xơ cũng làm tăng tiết axit mật trong đường tiêu hóa, gây ra sự hình thành các loại oxy phản ứng (ROS) và tổn thương DNA. ROS, chẳng hạn như hydrogen peroxide (H2O2), anion superoxide, và các gốc hydroxyl, được tạo ra trong các tế bào hiếu khí bằng cách khử không hoàn toàn oxy phân tử trong quá trình hô hấp của ty thể. Một số bệnh GI, bao gồm tổn thương niêm mạc, bệnh viêm ruột, loét dạ dày tá tràng và ung thư dạ dày cũng có liên quan đến sự mất cân bằng giữa việc hình thành ROS và khả năng chống oxy hóa.

Ăn nhiều rau, ngũ cốc và trái cây giàu chất chống oxy hóa có thể bảo vệ cơ thể để chống lại các bệnh mãn tính như tiểu đường, bệnh tim mạch và ung thư. Đặc biệt là các loại rau họ cải như bông cải xanh, bắp cải và súp lơ có chứa các hợp chất hoạt tính sinh học có đặc tính chống oxy hóa như isothiocyanates. Sulforaphane (SFN), một isothiocyanate có trong tự nhiên, đã được nghiên cứu về tác dụng bảo vệ tế bào và phòng ngừa hóa học. SFN không có tác dụng chống oxy hóa trực tiếp mà thay vào đó là chất hoạt hóa mạnh của yếu tố 2 liên quan đến erythroid hạt nhân 2 (NRF2), là chất chuyển đổi oxy hóa khử chủ. Sau khi kích hoạt, NRF2 cảm ứng sự biểu hiện của các enzym bao gồm thioredoxin reductase-1 (TXNRD1), heme oxygenase-1 (HMOX1), NAD (P) H: quinone oxidoreductase-1 (NQO1), và glutathione reductase (GSR) và từ đó có thể kích hoạt NRF2 để bảo vệ tế bào chống lại stress oxy hóa và chứng viêm.
Các nghiên cứu dược động học chỉ ra rằng tiêu thụ bông cải xanh trong chế độ ăn uống dẫn đến nồng độ đỉnh trong huyết tương SFN ∼2 μM. Nồng độ > 5 μM thường sẽ khó đạt được bằng chế độ ăn uống. Tác dụng bảo vệ tế bào qua trung gian NRF2 có thể đạt được ở nồng độ SFN thấp (<5 μM). Mặt khác, nồng độ SFN cao (> 20 μM) có thể gây ra quá trình apoptosis, ngừng chu kỳ tế bào và lão hóa ở các tế bào ung thư vú, tuyến tụy và dạ dày. Hoạt động ngăn ngừa hóa chất của SFN nồng độ cao đã được báo cáo ở một số loại tế bào ung thư khá phổ biến hiện nay.
Không có sự thống nhất nào liên quan đến độc tính của SFN trong các tế bào và mô của người bình thường. Một số nghiên cứu đã sử dụng SFN như một chất chống oxy hóa hoặc hợp chất gây độc tế bào mà không khám phá mối quan hệ giữa nồng độ và phản ứng. SFN có thể ít độc hơn đối với tế bào bình thường so với tế bào ung thư. Tuy nhiên, các tác động gây độc tế bào tương tự được quan sát thấy ở tế bào ung thư, chẳng hạn như tín hiệu apoptotic, bắt giữ chu kỳ tế bào và tổn thương DNA, cũng đã được quan sát thấy ở các tế bào bình thường. Xem xét rằng đường tiêu hóa là nguồn cung cấp ROS và SFN là chất chống oxy hóa, nghiên cứu này đã đánh giá tác động phân tử của SFN ở các nồng độ khác nhau trong dòng tế bào niêm mạc dạ dày của con người (MNP01). Bằng cách sử dụng các xét nghiệm dựa trên tế bào, nghiên cứu đã đánh giá tiềm năng gây độc tế bào và chống oxy hóa của SFN, và sử dụng giải trình tự thế hệ tiếp theo để kiểm tra những thay đổi trên biểu hiện gen.
2. Vật liệu và phương pháp
2.1. Thuốc thử nuôi cấy tế bào và điều kiện xử lý

DL-Sulforaphane, Triton ™ X-100, màu đỏ trung tính, 2 ′, 7′-dichlorofluorescin diacetate, và dimethyl sulfoxide (DMSO) được mua từ Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s Modified Eagle Medium với glucose cao (DMEM), penicillin-streptomycin 10.000 U / mL, và huyết thanh bò thai (FBS), được lấy từ Gibco (Carlsbad, CA, USA). MTT (3- (4,5-dimethylthiazol-2-yl) −2,5-diphenyltetrazolium bromide), StemPro ™ Accutase ™ Enzyme phân ly tế bào, TrypLE ™ Express Enzyme và p -nitrophenyl phosphate (pNPP) đã được mua từ Thermo Scientific (Công viên Canoga, CA, Hoa Kỳ).
Dòng tế bào niêm mạc dạ dày người (MNP01) được nuôi cấy trong 90% DMEM High Glucose bổ sung 1% penicillin-streptomycin và 10% FBS trong môi trường ẩm ướt ở 37°C với 5% CO 2. Tế bào MNP01 được cung cấp bởi Phòng thí nghiệm Di truyền Tế bào và Độc tính Di truyền Người của Đại học Liên bang Pará (Belém, Pará, Brazil). Môi trường nuôi cấy được thay đổi sau mỗi 2-3 ngày. Khi các mẫu cấy đạt đến sự hợp lưu, các tế bào được rửa hai lần bằng dung dịch muối đệm phosphat (PBS, pH 7,4), tách ra bằng TrypLE ™ và được nuôi cấy phụ. Tất cả các thí nghiệm đều được tiến hành giữa đoạn tế bào thứ ba và thứ tám. SFN được hòa tan trong DMSO, và các tế bào MNP01 cũng được xử lý bằng phương tiện kiểm soát (0,1% DMSO) và SFN, 2–32 μM.
2.2. Thử nghiệm khả năng sống của tế bào
Đối với ba thử nghiệm khả năng sống sót của tế bào, nghiên cứu tuân theo cùng một quy trình nuôi cấy và thu hoạch. Tế bào MNP01 được gieo vào đĩa 96 giếng, 1 × 10 4 mỗi giếng, và ủ ở 37°C, 5% CO2 trong 24 giờ (80–90% hợp lưu). Tế bào nuôi cấy được xử lý bằng SFN (2, 4, 8, 16, hoặc 32 μM) và kiểm soát phương tiện (0,1% DMSO). Sau khi xử lý 24 giờ, các giếng mẫu được rửa hai lần bằng PBS để bắt đầu các thử nghiệm khả năng sống, được thực hiện ba lần cho mỗi nhóm.
Thử nghiệm MTT được thực hiện theo quy trình của Mosmann. Dung dịch MTT (100 μL, 0,5 mg / mL) được thêm vào mỗi giếng, và đĩa được ủ trong tủ ấm CO 2 ở 37°C trong 3 giờ. Dung dịch MTT được loại bỏ và DMSO, 200 μL, được thêm vào để hòa tan các tinh thể formazan. Độ hấp thụ được đo ở bước sóng 570 nm bằng đầu đọc vi tấm ELx800 (Biotek-Winooski, VT, USA). Đối với xét nghiệm màu đỏ trung tính hấp thụ (NRU), dung dịch màu đỏ trung tính (100 μL, 40 μg / mL) được thêm vào mỗi giếng, và đĩa được ủ ở 37°C trong 3 giờ. Dung dịch giải hấp phụ màu đỏ trung tính (200 μL, 50 phần etanol, 49 phần nước khử ion, 1 phần axit axetic băng) được thêm vào mỗi giếng. Đĩa được lắc trong 30 phút và giữ yên trong 5 phút. Giá trị độ hấp thụ thu được ở bước sóng 540 nm bằng máy quang phổ iMark ™ (Bio-Rad, Hercules, CA, USA).
Hoạt động của acid phosphatase (APH) được xác định bằng cách sử dụng pNPP, như được Friedrich et al mô tả. Đệm xét nghiệm (100 μL, 0,1 M natri axetat, 0,1% Triton-X-100, bổ sung pNPP) và PBS, 100 μL, được thêm vào mỗi giếng, và đĩa được ủ ở 37°C trong 90 phút. Sau đó, NaOH (1 N, 10 μL) được thêm vào mỗi giếng và độ hấp thụ được đo ở bước sóng 405 nm bằng cách sử dụng đầu đọc vi tấm ELx800. Các chỉ số hấp thụ của các tế bào được xử lý được bình thường hóa so với các tế bào tiếp xúc với điều khiển phương tiện. Các giá trị được phân tích bằng ANOVA một chiều, sau đó là kiểm tra hậu Dunnett. Phân tích thống kê được thực hiện bằng GraphPad Prism 6 (San Diego, CA, USA).
2.3. Các loại oxy phản ứng
Sự sản sinh ROS được phát hiện bằng đầu dò huỳnh quang 2′, 7′-dichlorofluorescin diacetate (H 2 DCFDA), theo hướng dẫn của nhà sản xuất. Tế bào được cấy vào đĩa rắn đen 96 giếng, 1 × 10 4 mỗi giếng, và ủ ở 37°C, 5% CO 2 trong 24 giờ. Sau 12 và 24 giờ xử lý, phần nổi phía trên được loại bỏ và H2 DCFDA (10 μM) được thêm vào mỗi giếng, sau đó ủ 30 phút ở 37°C trong bóng tối.
Tiềm năng chống oxy hóa của SFN được đo bằng cách cho tế bào tiếp xúc với H2 O2 (0,2 mM) trong 30 phút, sau khi xử lý trước SFN (12 và 24 giờ). H2DCFDA được thêm vào mỗi giếng và ủ trong 30 phút ở 37°C trong bóng tối. ROS nội bào được đo bằng đầu đọc vi tấm ELx800 với bước sóng kích thích và phát xạ lần lượt là 485 và 528 nm.
2.4. Apoptosis: phân tích tế bào dòng chảy
Quá trình chết rụng tế bào được đo bằng phương pháp đo tế bào dòng chảy sử dụng phương pháp ghi nhãn Annexin V-FITC và propidium iodide (PI) (Thermo Fisher Scientific, Waltham, MA, USA). Các tế bào được xử lý bằng SFN, 0,1% DMSO (đối chứng âm tính), hoặc 1 μM doxorubicin (Fauldoxo, Libbs, Brazil) là đối chứng dương tính. Sau 24 giờ, tế bào được thu hoạch và rửa sạch bằng PBS, sau đó được ủ với Annexin V (0,5 μM) trong 15 phút trong bóng tối ở nhiệt độ phòng. Dung dịch PI (1 μg / mL) đã được thêm vào trước khi phân tích. Các tín hiệu Annexin V-FITC / PI được đánh giá bằng máy đo lưu lượng FACSCanto và phân tích bằng phần mềm FACSDiva (BD Biosciences, San Jose, CA, USA). Quần thể tế bào được đặt theo góc phần tư như sau:
- tế bào sống sót (Annexin V / PI kép âm tính)
- tế bào chết sớm (Annexin V dương tính / PI âm tính)
- tế bào chết muộn (Annexin V / PI kép âm tính)
2.5. Trích xuất RNA và chuẩn bị thư viện
Tế bào MNP01 được nuôi cấy và xử lý với 8 μM SFN trong 24 giờ. RNA tổng số được phân lập theo quy trình của nhà sản xuất PureLink® RNA Mini (Invitrogen; Carlsbad, CA, USA). RNA được định lượng bằng xét nghiệm Qubit 2.0 Phạm vi rộng (Invitrogen). Việc thu mRNA và xây dựng thư viện mRNA-Seq mắc kẹt được thực hiện bằng KAPA mRNA HyperPrep Kit (Roche; Basel, Switz.) trên Máy trạm xử lý chất lỏng tự động JANUS G3 (PerkinElmer; Waltham, MA, USA). Quy trình này bao gồm bắt giữ, rửa giải và phân mảnh mRNA, sau đó là tổng hợp sợi thứ nhất và sợi thứ hai. Các bộ điều hợp Illumina được gắn ở đầu của các đoạn cDNA có nguồn gốc mRNA, sau đó là quá trình khuếch đại PCR thư viện. Để kiểm tra chất lượng thư viện và sự hiện diện của sơn lót, hãy chạy trên 2200 TapeStation (Agilent; Santa Clara, CA,
2.6. Lập trình tự, ánh xạ và phân tích biểu thức vi phân
Các thư viện được giải trình tự trong một làn với 50 bp trên bộ giải trình tự Illumina HiSeq4000. Các lần đọc trình tự thô đã được kiểm tra chất lượng bằng FastQC trước quá trình ánh xạ. Bộ điều hợp và các lần đọc chất lượng thấp đã được cắt bỏ bằng cách sử dụng Trimmomatic phiên bản 0.38. Các lần đọc đã cắt bỏ qua bộ lọc chất lượng được ánh xạ tới chỉ số bộ gen tham chiếu ( Homo sapiens Ensembl GRCh38, phiên bản 90) bằng cách sử dụng công cụ căn chỉnh STAR. Số lần đọc cho mỗi gen được tính toán cho mỗi mẫu. Các gen có số lượng bằng 0 trong tất cả các mẫu đã được lọc ra khỏi ma trận số lượng gen. Các mẫu xử lý được kiểm tra dựa trên các mẫu đối chứng để tìm biểu hiện khác biệt bằng cách sử dụng gói Bioconductor DESeq2. Danh sách gen biểu hiện khác biệt được lọc bằng cách sử dụng tỷ lệ phát hiện sai 1% (FDR) và thay đổi nếp gấp tuyệt đối > 2 đối với các gen điều chỉnh lên và gen điều chỉnh giảm. Danh sách gen được tách thành gen điều chỉnh lên và gen gen điều chỉnh giảm. Phân tích đại diện của con đường được xác định bằng cách sử dụng ConsensusPathDB. Cơ sở dữ liệu KEGG, Reactome và WikiPathways đã được sử dụng và sự chồng chéo số gen tối thiểu được thiết lập lên đến 2 với giới hạn giá trị p là 0,01. Phân tích bản thể gen (GO) mỏng được thực hiện bằng cách sử dụng WebGestAlt với quy trình FDR 1% và Benjamini-Hochberg làm điều chỉnh nhiều thử nghiệm đã chọn. Việc giàu GO của các quá trình sinh học và phân tích các nhiễu loạn về hóa học và di truyền được thực hiện bằng phần mềm GSEA 4.03 với ngưỡng cắt 1% FDR và bộ gen có kích thước tối thiểu là 15 bằng cách sử dụng cơ sở dữ liệu ký hiệu phân tử (MSigDB). Mạng lưới các cụm GO được bổ sung phong phú từ bộ dữ liệu được kiểm soát lên và xuống được tạo bởi Cytoscape phiên bản 3.7.2 với FDR 1% và ngưỡng tương tự ≥ 0,5. Bản đồ nhiệt được tạo với các tệp kết quả từ GSEA bằng cách sử dụng Gitools 2.3.1.
3. Các kết quả thu được từ nghiên cứu
3.1. Độc tính tế bào
Tế bào MNP01 được xử lý bằng SFN (2–32 μM), hoặc DMSO (đối chứng) trong 24 giờ. Độc tính tế bào được đánh giá bằng ba xét nghiệm so màu khác nhau: MTT, APH và NRU (Hình 1). SFN không làm giảm khả năng sống của tế bào ở 2 hoặc 4 μM, nhưng SFN gây độc tế bào ở 8–32 μM (xét nghiệm MTT và APH) và 16 và 32 μM (xét nghiệm NRU).

Hình 1. Khả năng sống của tế bào của MNP01 được xử lý bằng sulforaphane sử dụng các xét nghiệm so màu khác nhau: (A) MTT, (B) Hoạt tính của acid phosphatase, và (C) Xét nghiệm màu đỏ trung tính sau 24 giờ. Các giá trị được đánh dấu bằng (các) dấu hoa thị là khác biệt đáng kể (* P <0,05, ** P <0,01 và *** P <0,001) so với Đối chứng (DMSO 0,1%) sau ANOVA sau đó là thử nghiệm của Dunnett. Các kết quả được trình bày dưới dạng giá trị trung bình ± độ lệch chuẩn từ ba thí nghiệm ba lần độc lập (n = 9).
3.2. Tác dụng bảo vệ của SFN chống lại sự hình thành ROS do H2O2 gây ra
Nghiên cứu đo lường ROS bằng đầu dò H2 DCFDA. Sau 12 giờ xử lý, chỉ có 32 μM SFN làm giảm sự hình thành ROS (Hình 2 A). Tuy nhiên, sau 24 giờ xử lý, SFN làm giảm ROS ở tất cả các nồng độ được thử nghiệm (Hình 2B). Các tế bào được xử lý SFN tạo ra mức ROS thấp hơn so với các tế bào kiểm soát sau 12 hoặc 24 giờ điều trị (Hình 2 C, D).

Hình 2. Xét nghiệm sự hình thành ROS nội bào trong các tế bào MNP01 sau (A) 12 giờ và (B) 24 giờ điều trị với SFN. Mức ROS sau (C) 12 giờ và (D) 24 giờ điều trị với SFN, sau đó tiếp xúc với H 2 O 2 (0,2 mM) trong 30 phút. Các giá trị được đánh dấu bằng (các) dấu hoa thị là khác biệt đáng kể (* P <0,05, ** P <0,01 và *** P <0,001) so với Đối chứng (DMSO 0,1%) sau ANOVA sau đó là thử nghiệm của Dunnett. Mức ROS được biểu thị dưới dạng đơn vị huỳnh quang tùy ý. Các kết quả được trình bày dưới dạng giá trị trung bình ± SD từ ba thí nghiệm ba lần độc lập (n = 9).
3.3. Sự chết tế bào
Apoptosis được đo bằng cách ghi nhãn, phosphatidylserine tiếp xúc được đo bằng cách sử dụng ghi nhãn Annexin V và PI đo bằng phép đo tế bào dòng chảy. Apoptosis tăng đáng kể ở nồng độ SFN cao (16 và 32 μM; Hình 3 A, B), làm giảm khả năng sống của tế bào ở các nồng độ này (Hình 3 C).

Hình 3. Đo độ chết của MNP01 được điều trị bằng sulforaphane trong 24 giờ. (A) Kết quả đo lưu lượng tế bào với ghi nhãn Annexin V và propidium iodide (PI). Tế bào sống được (Q3): Annexin V và PI kép âm tính; Tế bào Apoptotic sớm (Q4): Annexin V dương tính và PI âm tính; Tế bào chết / hoại tử muộn (Q2): Annexin V và PI dương tính kép; Tế bào chết (Q1): Annexin V âm tính và PI dương tính. Biểu đồ thanh thể hiện phần trăm (B) tế bào chết / hoại tử và (C) tế bào sống sót sau khi điều trị bằng sulforaphane. Các giá trị được đánh dấu bằng (các) dấu hoa thị là khác biệt đáng kể (** P <0,01 và *** P <0,001) so với Đối chứng (DMSO 0,1%) sau ANOVA sau đó là thử nghiệm của Dunnett. Dữ liệu đại diện cho giá trị trung bình ± SD từ ba thí nghiệm độc lập (n = 3).
3.4. Hồ sơ phiên mã
Nghiên cứu đã chọn một nồng độ trung gian để khảo sát ảnh hưởng của SFN đối với sự biểu hiện gen. SFN, 8 μM, cho thấy tác dụng chống oxy hóa và bảo vệ tế bào tương tự ở nồng độ thấp hơn (2 và 4 μM), và mặc dù nó là chất độc tế bào, nhưng khả năng tồn tại không giảm đáng kể. Cấu hình RNA-Seq cho thấy 409 gen biểu hiện khác biệt (DEG), 280 gen quy định lên và 129 gen quy định giảm. Công cụ trực tuyến “Webgestalt” đã được sử dụng để điều tra các quá trình sinh học, các thành phần tế bào và chức năng phân tử của chúng.
“Quy định sinh học” là cụm GO thường xuyên nhất trong danh mục “quá trình sinh học”. Các loại “thành phần tế bào” và “chức năng phân tử” phổ biến nhất lần lượt là liên kết màng và protein (Hình 4A). Một bản đồ nhiệt của các DEG được điều chỉnh tăng và giảm nhất được theo sau bởi phân tích giàu lộ trình cho thấy một số DEG được điều chỉnh tăng liên quan đến các đặc tính chống oxy hóa của SFN (Hình 4B). Các gen trong con đường chuyển hóa glutathione, stress oxy hóa và NRF2 được giàu trong số các DEG được điều chỉnh, bao gồm GSR , HMOX1 và TXNRD1, mã hóa các enzym chống oxy hóa. Các DEG được điều chỉnh cao cũng tham gia vào lộ trình tín hiệu p53, bao gồm CDKN1A , MDM2 và BBC3.

Hình 4. Phân tích làm giàu chức năng của các gen biểu hiện khác biệt trong các tế bào MNP01 được xử lý bằng sulforaphane (SFN 8 μM) trong 24 giờ. (A) Phân loại chức năng bản thể học gen của tất cả các gen biểu hiện khác biệt. (B) Bản đồ nhiệt đại diện cho cấu hình biểu hiện gen và các con đường làm giàu nhất dựa trên giá trị q. Bản đồ nhiệt minh họa mức độ biểu hiện gen từ thấp hơn (xanh lam) đến cao hơn (đỏ). Mỗi cột đại diện cho một bản sao sinh học của các mẫu SFN so với đối chứng.
Để mô tả thêm các chức năng sinh học của DEG, nghiên cứu đã thực hiện phân tích giàu chuyên sâu với MSigDB, sử dụng quy trình sinh học GO và các rối loạn về di truyền và hóa học với GSEA (Hình 5). cụm phong phú nhất cho quá trình sinh học GO có liên quan đến phản ứng viêm (Hình 5 A). Nghiên cứu cũng xác định sự giàu cho các cụm của stress oxy hóa, chẳng hạn như ‘Giải độc’ và ‘Phản ứng với stress oxy hóa’, cả hai đều liên quan đến con đường NRF2, một mục tiêu nổi tiếng của SFN, xác thực cách tiếp cận của nghiên cứu. Các DEG được điều chỉnh tối đa cũng đã làm phong phú thêm cụm ‘Quy định về lộ trình tín hiệu apoptotic bên ngoài’. Đối với các DEG được điều chỉnh thấp, các cụm GO phong phú nhất có liên quan đến chu kỳ tế bào, chẳng hạn như ‘sao chép DNA’ và ‘phân bào giảm phân’. Chúng tôi cũng tìm thấy các cụm liên quan đến việc sửa chữa và hư hỏng DNA, chẳng hạn như ‘Sửa chữa đứt gãy sợi đôi và’ sửa chữa DNA’ (Hình 5 A).

Hình 5. Phân tích bản thể học gen của các gen biểu hiện khác biệt trong các tế bào MNP01 được điều trị bằng sulforaphane (8 μM) trong 24 giờ. (A) Bản đồ nhiệt đại diện cho các thuật ngữ quá trình sinh học được làm giàu của các gen biểu hiện khác biệt. (B) Bản đồ nhiệt đại diện cho các thuật ngữ được bổ sung thêm từ Lo lắng về hóa học và di truyền. Mạng lưới các quá trình sinh học được tạo ra bởi Cytoscape (1% FDR và độ tương đồng ≥ 0,5) của các thuật ngữ đã được làm giàu của các gen (C) được điều chỉnh tăng và (D) được biểu hiện khác biệt. Kích thước nút tương ứng với số lượng gen biểu hiện khác biệt trong thuật ngữ bản thể học gen.
Bằng cách sử dụng MSigDB Hóa chất và Rối loạn Di truyền, nghiên cứu đã xác định các dấu hiệu DEG bị thay đổi bởi SFN tương tự như các nhiễu loạn hóa học và di truyền khác đã được mô tả (Hình 5 B). Ví dụ, các gen điều chỉnh giảm được tìm thấy, trong một nghiên cứu khác, có liên quan đến sự chuyển đổi biểu mô-trung mô và di căn trong ung thư vú. Hơn nữa, các DEG điều chỉnh giảm tương tự như một cụm gen điều chỉnh lên liên quan đến sự tăng sinh tế bào do HPV gây ra và kiểu hình xâm lấn trong ung thư biểu mô cổ tử cung.
Nghiên cứu cũng đã thực hiện phân tích độ tương tự với các cụm được giàu bằng cách sử dụng Cytoscape và các mạng cho các cụm được giàu DEG được điều chỉnh lên và xuống đã được tạo ra. Các cụm giàu DEG được điều chỉnh tối đa hình thành hai mạng lưới nhỏ liên quan đến stress oxy hóa và phản ứng viêm ( Hình 5 C). Các cụm giàu DEG được điều chỉnh giảm tạo ra hai mạng lưới phức tạp liên quan đến chu trình tế bào và sửa chữa sao chép DNA ( Hình 5 D).
4. Thảo luận sau nghiên cứu

Đường tiêu hóa là nguồn cung cấp ROS và cũng là nơi rất nhạy cảm với tổn thương do oxy hóa. Bất chấp hàng rào biểu mô, một số bệnh lý GI gây ra do mất cân bằng sản xuất ROS. Chất chống oxy hóa có thể hạn chế tác động bất lợi của ROS bằng cách loại bỏ các gốc tự do và ức chế quá trình oxy hóa. SFN, một isothiocyanate được tìm thấy trong các loại rau họ cải, là một hợp chất hoạt tính sinh học kích hoạt NRF2 và gián tiếp có thể hoạt động như một chất chống oxy hóa.
Việc kích hoạt các enzym liên hợp xenobiotic bằng SFN có thể có tác dụng bảo vệ tế bào trong các dòng tế bào và trong các nghiên cứu trên động vật. Tác dụng bảo vệ của SFN chống lại stress oxy hóa trong tế bào GI đã được quan sát thấy ở những con chuột bị nhiễm vi khuẩn Helicobacter pylori và trong các tế bào nuôi cấy. Kết quả của chúng tôi cho thấy SFN không tạo ra ROS; thay vào đó, xử lý SFN dẫn đến giảm sản xuất ROS và ứng suất oxy hóa gây ra bởi H 2O2 phơi bày. Tuy nhiên, ít tế bào có thể sống được ở nồng độ SFN cao hơn. Phân tích cho thấy lộ trình NRF2 là quan trọng nhất trong số các DEG được điều chỉnh tăng và phân tích GO cho thấy các cụm giàu DEG đã được điều chỉnh liên quan đến phản ứng stress oxy hóa. Dữ liệu RNA-Seq chỉ ra rằng xử lý bằng SFN có điều chỉnh một số enzym chống oxy hóa, chẳng hạn như HMOX1, NQO1 và TXNRD1. Một nghiên cứu trước đây đã chỉ ra rằng sự điều hòa HMOX1 và NQO1 trong các tế bào đại tràng sơ cấp của con người được điều trị bằng SFN thực hiện hoạt động bảo vệ tế bào bằng cách giải độc xenobiotics oxy hóa. Một nghiên cứu khác đề xuất vai trò bảo vệ của SFN trong việc ngăn chặn sự hình thành ROS qua trung gian cảm ứng HMOX1.
Nghiên cứu này cũng tìm thấy các gen được điều chỉnh liên quan đến quá trình chuyển hóa glutathione, điều này vô cùng hứa hẹn vì SFN được liên hợp nhanh chóng với glutathione. GCLC và GCLM là các gen mã hóa các tiểu đơn vị của glutamate-cysteine ligase, một loại enzyme sinh tổng hợp glutathione. Một số nghiên cứu cho thấy vai trò bảo vệ của SFN chống lại stress oxy hóa bằng cách điều chỉnh tăng GCLC và GCLM. Trong các tế bào tủy xương của con người tiếp xúc với H2O2, tiền xử lý với SFN đã ngăn chặn sự mất khả năng sống và bảo vệ ty thể bằng cách tăng cường mức glutathione. Kết quả của chúng tôi xác nhận rằng SFN ở nồng độ thấp làm tăng sự biểu hiện của các gen chống oxy hóa.
SFN có thể gây độc tế bào. Các đặc tính bảo vệ của SFN, bao gồm apoptosis, bắt giữ chu kỳ tế bào và tổn thương DNA, phụ thuộc vào liều lượng / nồng độ. Ví dụ, các tế bào ung thư dạ dày cho thấy khả năng tồn tại giảm đáng kể ở 5–20 μM. Hầu hết các tài liệu về tác dụng gây độc tế bào của SFN tập trung vào các tế bào ung thư. Tuy nhiên, nồng độ SFN cao cũng gây độc cho các tế bào không phải ung thư. Tác dụng ngoài ý muốn của SFN nồng độ cao đã được báo cáo. Ví dụ, 20 μM SFN gây ra đứt sợi đơn trong tế bào nội mô tĩnh mạch rốn của con người. Một nghiên cứu trên tế bào gốc trung mô cho thấy SFN, ở nồng độ thấp, bảo vệ tế bào chống lại stress oxy hóa gây ra. Mặt khác, nồng độ SFN cao gây ra độc tính tế bào và tổn thương DNA.
SFN có thể gây ra thiệt hại cho ty thể, ví dụ, phá vỡ điện thế màng ty thể, có thể dẫn đến sự biểu hiện của các yếu tố pro-apoptotic, chẳng hạn như protein tế bào B lymphoma 2 (BCL2). Mặt khác, nồng độ SFN thấp có thể bảo vệ ti thể. Chúng tôi đã thấy sự khác biệt giữa kết quả xét nghiệm MTT và APH so với xét nghiệm NRU. Trong một nghiên cứu gần đây, các tế bào HeLa được xử lý với 10 μM SFN cho thấy chức năng / thế hệ lysosome tăng lên. Quá trình sinh sinh lysosome góp phần vào việc thanh thải hiệu quả các ty thể bị rối loạn chức năng. Cơ chế này gợi ý một lời giải thích khả dĩ cho kết quả xét nghiệm NRU của chúng tôi rằng 8 μM SFN không làm thay đổi khả năng sống của tế bào.
Chúng tôi quan sát thấy rằng các tế bào được điều trị bằng SFN chia sẻ các DEG tương tự với các bộ dữ liệu về hóa học và di truyền khác liên quan đến cảm ứng apoptosis. Ví dụ, một nghiên cứu với tế bào NT2 / D1 được điều trị bằng cisplatin và một nghiên cứu khác nghiên cứu tác động của epoxit trong tế bào SH-SY5Y cho thấy cơ chế tự chết tương tác. Một trong những gen được chia sẻ trong phân tích Lo lắng về hóa học và di truyền là FAS, gen này chịu trách nhiệm mã hóa các thụ thể chết trong con đường tự chết bên ngoài.
SFN gây ra sự điều hòa của các gen tín hiệu p53. Mạng lưới tín hiệu này kiểm soát nhiều phản ứng của tế bào, chẳng hạn như bắt giữ chu kỳ tế bào, sửa chữa DNA và lão hóa do các kích thích căng thẳng, ví dụ, tổn thương DNA. Trong dữ liệu của chúng tôi, CDKN1A và các gen phản ứng tổn thương DNA khác đã được SFN điều chỉnh. CDKN1A đóng một vai trò quan trọng trong việc điều chỉnh sự tiến triển của chu kỳ tế bào; protein được mã hóa làm trung gian bắt giữ chu kỳ tế bào phụ thuộc p53 để đáp ứng với các kích thích căng thẳng. Chúng tôi cũng quan sát thấy sự phong phú của một số lượng lớn các cụm GO trong chu kỳ tế bào, là kết quả của việc SFN điều chỉnh giảm một số gen liên quan đến chu kỳ tế bào. Hầu hết các gen này chịu trách nhiệm sửa chữa, sao chép và tổng hợp DNA, chẳng hạn như POLE2 ,E2F2 và DNA2. Chúng tôi cũng quan sát thấy sự điều hòa giảm của một số gen chịu trách nhiệm phân ly nhiễm sắc thể, chẳng hạn như CDCA5 và SPC24. Một số nghiên cứu đã báo cáo những bất thường về sự phân bào do SFN gây ra trong các bệnh ung thư, bao gồm ung thư dạ dày.
Tóm lại, kết quả của chúng tôi cho thấy nồng độ SFN thấp bảo vệ chống lại stress oxy hóa và sự hình thành ROS mà không có tác dụng gây độc tế bào. Những nồng độ này gần với mức huyết tương có thể đạt được bằng cách uống bông cải xanh hoặc các loại rau họ cải khác, ∼2 μM. Tuy nhiên, SFN, 8 μM gây ra sự điều hòa của các cơ quan điều hòa apoptotic tích cực, điều hòa giảm các gen liên quan đến sửa chữa tổn thương DNA và kiểm soát chu kỳ tế bào, và điều chỉnh sự biểu hiện gen của tế bào MNP01 thành kiểu hình pro-apoptotic, đồng thời phát huy đặc tính chống oxy hóa của nó, thể hiện qua việc điều chỉnh các gen liên quan đến các enzym chống oxy hóa. Nồng độ SFN cao có thể gây ra biểu hiện gen đáp ứng tổn thương DNA và các bất thường về chu kỳ tế bào có thể dẫn đến quá trình apoptosis. Mô tả sâu hơn về các hiệu ứng SFN, chẳng hạn như nghiên cứu động học và động lực học độc tính, sẽ làm rõ hiểu biết của chúng ta về tính an toàn và độc tính của hợp chất hoạt tính sinh học này.
4. Các chủ đề liên quan
Dưới đây là một số bài viết về các chủ đề nghiên cứu có liên quan đến hoạt chất Sulforaphane và công dụng của chúng:
- Mầm bông cải xanh giàu chất cảm ứng của các enzyme bảo vệ giúp chống lại ung thư
- Tác dụng bảo vệ của chiết xuất bông cải xanh và Sulforaphane chống lại sự căng thẳng oxy hóa
- Sulforaphane : Một sản phẩm tự nhiên chống lại các loại oxy phản ứng
5. Tham vấn chuyên môn
- Bác sĩ
- Nguồn tham khảo: https://www.sciencedirect.com/science/article/pii/S1383571820300711